


Navigating gut-liver metabolic homeostasis: the role of the autonomic nervous system
 0
0 
Abstract
The frequent clinical co-occurrence of intestinal and hepatic disorders points to underlying bidirectional communication along the gut-liver axis. Existing research has primarily focused on metabolic and humoral interactions, including bile acid circulation and portal venous signaling, whereas the autonomic nervous system (ANS) has received comparatively limited attention as an integrative regulatory pathway. This review reframes the axis through a neural perspective, arguing that the ANS functions as a master coordinator of metabolic homeostasis, linking immune regulation, metabolic function, and gut microbiota activity into a unified neuro-immune-endocrine network. The sympathetic and parasympathetic branches, along with the enteric nervous system, modulate barrier integrity, immune cell trafficking, hepatic glucose and lipid metabolism, and microbial composition via neurotransmitter-mediated and receptor-dependent signaling pathways. Dysregulation of this autonomic circuitry disrupts metabolic balance and is implicated in the shared pathophysiology of conditions such as inflammatory bowel disease (IBD) and metabolic dysfunction-associated steatotic liver disease (MASLD). Conversely, neuromodulation strategies, particularly noninvasive vagus nerve stimulation, have demonstrated therapeutic potential in reducing inflammation and restoring metabolic balance. By positioning the ANS as one of the central regulators of gut-liver metabolic cross-talk, this review advances current mechanistic understanding of gut-liver comorbidities and highlights bioelectronic medicine as a promising therapeutic strategy for metabolic disorders. This perspective bridges neuroscience, gastroenterology, and hepatology, offering novel insights for the management of complex, multi-organ metabolic diseases.

Keywords
INTRODUCTION
Intestinal and hepatic diseases represent a growing global health burden due to their rising incidence and substantial clinical impact[1,2]. For instance, the worldwide prevalence of inflammatory bowel disease (IBD) increased by 88.3% from 1990 to 2021[3]. Similarly, the epidemiological footprint of metabolic dysfunction-associated steatotic liver disease (MASLD) has escalated dramatically, with recorded cases surging from 78,082 in 2010 to 621,400 in 2021[4]. In parallel, a growing body of literature underscores a robust, bidirectional cross-talk between the intestine and liver[5]. While a high-fat diet predisposes preclinical models to colitis[6,7], patients with IBD exhibit an elevated risk of developing MASLD[8,9]. As the prevalence of these metabolic and inflammatory conditions continues to rise in parallel with industrialization and lifestyle changes, it is imperative to identify key contributing factors in modern high-paced societies.
Dietary imbalance and overnutrition are well-recognized contributors to intestinal and hepatic dysfunction through their effects on lipid accumulation and systemic metabolic dysregulation. More recently, accelerated societal cadences and escalating psychosocial stress have emerged as formidable disruptors of metabolic homeostasis, particularly in the intestine and liver. Psychological stress influences not only endocrine pathways involved in metabolic balance but also the autonomic nervous system (ANS), which coordinates metabolic homeostasis, enteric nervous activity, immune responses, and rapid stress adaptation in these organs[10].
Composed of the sympathetic, parasympathetic, and enteric divisions, the ANS orchestrates metabolic homeostasis, immune regulation, and stress adaptation through neurotransmitter release and neural circuits[11-13]. Growing evidence indicates that ANS dysfunction is closely associated with a high incidence of gastrointestinal and hepatic diseases, particularly those involving metabolic components[14]. Notably, intestinal and hepatic disorders frequently co-occur with psychological disturbances such as anxiety and depression, and emotional states are known to influence disease progression in conditions like irritable bowel syndrome and MASLD. Anatomically, the ANS establishes a direct signaling pathway between the gut and liver. The vagus nerve originates from the medulla oblongata and extensively innervates both organs, while shared brainstem nuclei integrate hepatic and intestinal neural inputs. Together, these neural connections provide a structural basis for inter-organ communication and metabolic coordination[3,15-18]. Collectively, these observations suggest that ANS dysregulation may play a critical role in the pathogenesis of gut-liver metabolic comorbidities[19]. Nevertheless, the granular neural circuitries whereby the ANS orchestrates this inter-organ metabolic dialogue remain largely enigmatic, thereby warranting a systematic synthesis.
In this review, we summarize current clinical and preclinical evidence regarding the regulatory roles of the ANS in intestinal and hepatic physiology and pathology, with particular emphasis on metabolic regulation. We further discuss emerging bioelectronic neuromodulation strategies and their effects on the gut microbiome and hepatic metabolism, providing a translational framework that links basic neuroscience with clinical gastroenterology and hepatology.
THE ANS IN THE GASTROINTESTINAL METABOLIC REGULATION
Distribution of the ANS in the gastrointestinal tract
The ANS functions as a major neural interface between the central nervous system (CNS) and the gut. The gut nervous system comprises the enteric nervous system (ENS), sympathetic nervous system (SNS), and parasympathetic nervous system (PNS)[20,21]. The ENS, an intrinsic neural network within the gastrointestinal tract, comprises a meshwork of enteric neurons and glial cells embedded within the myenteric and submucosal plexuses. It senses intestinal alterations and maintains normal gastrointestinal function[22]. The SNS and PNS belong to the exogenous nervous system. Preganglionic sympathetic neurons innervating the upper gastrointestinal tract arise from the T6-T9 thoracic spinal segments, while those destined for the colon originate within the L2-L5 lumbar spinal segments [Figure 1][23]. Most postganglionic sympathetic neurons innervating the gastrointestinal tract are located within the prevertebral ganglia, with some originating from paravertebral ganglia[24,25]. The intestinal parasympathetic nerves consist of the vagus nerve and sacral plexus nerves[26]. The vagus nerve, which provides widespread innervation to the stomach, small intestine, and proximal colon, arises from a discrete topographic network within the brainstem, encompassing the dorsal motor nucleus of the vagus, the nucleus tractus solitarius, and the nucleus ambiguus. Sacral plexus nerves in the distal colon originate primarily from preganglionic neurons within the S1-S4 spinal cord[27,28]. Unlike the SNS, the PNS lacks a continuous ganglion chain. Instead, it comprises discrete ganglia juxtaposed to their terminal target organs, featuring elongated preganglionic fibers coupled with shortened postganglionic counterparts[29].
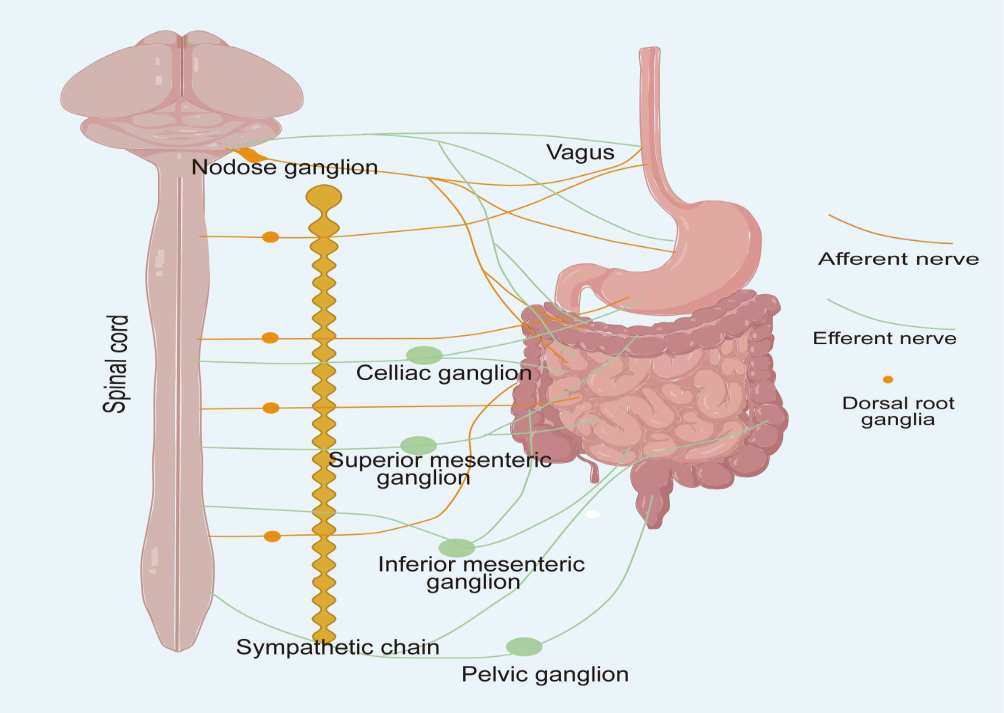
Figure 1. Sympathetic input to the gastrointestinal tract is conveyed by postganglionic efferent neurons that arise from the celiac, superior mesenteric, and inferior mesenteric ganglia. Sympathetic fibers projecting to the stomach, duodenum, and small intestine originate from spinal segments T6-T9, whereas those innervating the colon derive from segments L2-L5. Parasympathetic innervation is provided by the vagus nerve and pelvic splanchnic nerves. The cell bodies of vagal sensory neurons are located in the nodose ganglion. In contrast, general visceral afferent fibers, including those associated with sympathetic and pelvic splanchnic pathways, travel alongside their corresponding efferent nerves, and their neuronal cell bodies are found in the dorsal root ganglia. Image created with Adobe Illustrator.
Autonomic regulation of intestinal metabolic functions
ANS is involved in coordinating and regulating key functions of the gastrointestinal tract via neurotransmitter release and neural circuits [Table 1], including movement, secretion, vasomotor regulation and local reflexes, all of which fundamentally sustain efficient nutrient absorption and metabolic processing. Both the small and large intestines possess considerable autonomous control and can function independently without exogenous neural input[30]. This intrinsic activity is primarily mediated by the ENS through the pacemaker function of interstitial cells of Cajal. The ENS modulates gastrointestinal function through neuropeptides such as vasoactive intestinal peptide and cholecystokinin, contributing to digestive function[31]. The coordinated action of the PNS and SNS, which encompasses motility, vasoconstriction/vasodilation, and glandular secretion, provides finer control over gastrointestinal function than either system alone, and all these processes are critical for optimal nutrient metabolism and energy balance. Overall, maintenance of autonomic homeostasis is essential for normal intestinal physiology and systemic metabolic balance.
The primary functions of key neurotransmitter in the gut-liver axis
| Neurotransmitters | Receptors | Functions | |
| SNS | NE | β2-AR | Increased hepatic glucose output; Inhibition of tight junction protein expression; Involvement in inflammatory responses; Regulation of tumor metabolism; Contraction of intestinal smooth muscle |
| β3-AR | Attenuation of cAMP signaling and promotion of lipid synthesis in hepatocytes | ||
| α1-AR | Contraction of intestinal smooth muscle | ||
| PNS | ACh | mAChR | Relaxation of intestinal smooth muscle; Promotion of bile acid secretion; Promotion of digestive juice secretion |
| nAChR | Anti-inflammatory response | ||
| VIP | VPAC2 | Relaxation of intestinal smooth muscle | |
| ENS | VIP | VPAC1 | Regulation of intestinal stem cell differentiation and relaxation of intestinal smooth muscle |
| VPAC2 | Contraction of intestinal smooth muscle | ||
| CCK | CCK1R | Regulation of intestinal peristalsis; Stimulation of gallbladder contraction; Participation in satiety signaling |
ANS regulates gastrointestinal motility
The movement of the gastrointestinal tract is coordinated by the ENS, SNS and PNS to ensure proper nutrient digestion and absorption. As a general paradigm, parasympathetic inputs predominantly exert pro-kinetic, excitatory influences, whereas sympathetic signaling operates as an overarching inhibitory brake. In the upper gastrointestinal tract, the PNS achieves precise regulation of gastrointestinal motility through two different signal inputs. The excitatory cholinergic pathway drives smooth muscle contraction via the activation of muscarinic cholinergic receptors on gastrointestinal musculature, whereas the nonadrenergic, noncholinergic pathway causes smooth muscle relaxation through mainly the release of nitric oxide and/or vasoactive intestinal polypeptide[32]. Regulation of this signaling is achieved by ENS integration and transmission of command signals to the myenteric plexus, rather than direct input to smooth muscle[33]. The inhibitory effect of SNS on gastrointestinal motility is likewise indirect, mediated by the provision of inhibitory presynaptic inputs that constrain enteric neuronal excitation[34]. It has been suggested that sympathetic axons preferentially target cholinergic excitatory neurons rather than nitrergic neurons, reducing the release of excitatory neurotransmitters and suppressing intestinal motility[35]. Selective modulation of specific neural signaling pathways enables precise regulation of intestinal peristalsis and gastrointestinal metabolic function.
ANS regulates the blood flow in the gastrointestinal tract
The regulation of blood flow in the gastrointestinal tract is under bidirectional control by the ANS, with sympathetic nerves primarily mediating vasoconstriction and parasympathetic nerves promoting vasodilation. This mechanism ensures adequate delivery of nutrients and metabolic substrates. Nucleus tractus solitarius (NTS) receives vagal afferent input and provides strong excitatory projections to neurons in the rostral ventrolateral medulla (RVLM) which is a region containing sympathetic premotor neurons[36,37]. These RVLM neurons project directly to sympathetic preganglionic neurons located in the intermediolateral cell column of the spinal cord and modulate visceral blood flow and metabolic substrate distribution. The vagus nerve responds to diverse mechanical, chemical, and osmotic stimuli, including low-intensity gastric distension and bitter substances that activate RVLM neurons[38]. In contrast, systemic administration of cholecystokinin (CCK) suppresses the activity of RVLM neurons. CCK released from enteroendocrine cells (EECs) in response to food intake not only induces gastrointestinal relaxation via the vagus nerve but also regulates postprandial splanchnic blood flow through a vagal-sympathetic vasomotor reflex. These coordinated responses help maintain optimal conditions for postprandial metabolism[39,40].
ANS regulates the secretion of glands in the gastrointestinal tract
Intestinal gland secretion is regulated by intricate neural mechanisms, wherein the ENS, SNS, and PNS each play important roles in coordinating digestive enzyme and hormone secretion to maintain proper nutrient metabolism. The ENS regulates the proliferation and differentiation of intestinal stem cells through the release of vasoactive intestinal peptide (VIP)[41], dictating both the density and secretory repertoire of specialized mucosal cells, including enterochromaffin populations[42]. The synergistic interaction between the SNS and PNS is evident in the control of various digestive secretions. For instance, in salivary secretion, parasympathetic fibers via the facial and glossopharyngeal nerves directly stimulate salivary secretion to initiate carbohydrate digestion, while sympathetic fibers primarily induce contraction of myoepithelial cells to facilitate saliva expulsion[20]. Similar synergistic mechanisms govern the secretion of gastric juice, pancreatic juice, and bile[20]. The SNS and PNS coordinately regulate digestive gland output in response to physiological demands, supporting efficient nutrient processing and metabolic homeostasis.
THE ANS IN HEPATIC METABOLIC CONTROL
Distribution of the ANS in the liver
Sympathetic nerves to the liver originate from the thoracolumbar spinal cord (T7-T12). Preganglionic fibers travel by means of greater splanchnic nerves to the celiac and superior mesenteric ganglia, where synapses form[43]. Postganglionic fibers, primarily releasing norepinephrine (NE), then innervate the liver[43,44]. Parasympathetic innervation primarily originates fromthe craniosacral region, predominantly cholinergic (acetylcholine, ACh), and mainly via the hepatic branches of the vagus nerve, exerting a stimulatory effect on liver function[43,45,46]. Both sympathetic and parasympathetic fibers enter the liver under the mediation of hepatic portal, forming plexuses around three major vascular structures: the hepatic artery, portal vein, and bile duct[47]. The anterior hepatic plexus primarily distributes along the hepatic artery, while the posterior hepatic plexus distributes along the extrahepatic bile duct and portal vein[44-47] [Figure 2].
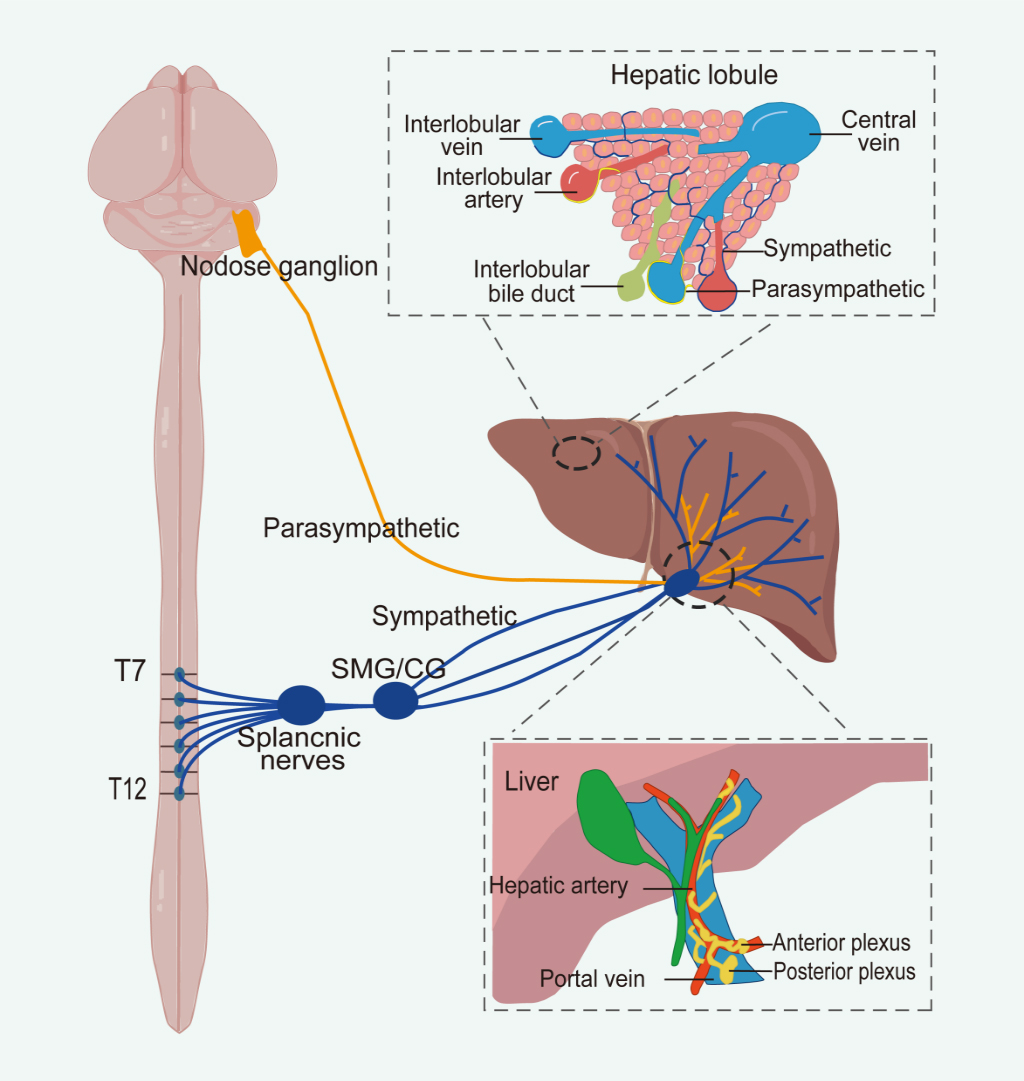
Figure 2. The sympathetic innervation of the liver arises from spinal segments T7 to T12, whereas parasympathetic innervation originates from nodose ganglion within the craniosacral regions. At the hepatic hilum, both sympathetic and parasympathetic fibers contribute to the formation of anterior and posterior autonomic plexuses. The anterior plexus surrounds the common hepatic artery, while the posterior plexus is located along the portal vein. Within the liver parenchyma, autonomic nerves are distributed around the hepatic artery, portal vein, and bile ducts. Notably, sympathetic fibers extend through the connective tissue and penetrate into the hepatic lobules, ultimately reaching the hepatocytes. Image created with Adobe Illustrator. SMG: Superior mesenteric ganglion; CG: celiac ganglion.
Autonomic regulation of hepatic metabolic functions
The ANS is a pivotal contributor to regulating liver metabolism, such as hepatic glucose, lipid, and bile acid metabolism, making the liver a central hub for systemic metabolic control.
Rapid regulation of glucose homeostasis is mediated by hepatic autonomic efferent pathways. Sympathetic nerve terminals release NE, thereby engaging hepatocyte β2-adrenergic receptors (β2-ARs) to augment hepatic glucose output[48,49]. Neuropeptides like galanin released from SNS terminals can also enhance hepatic glucose metabolism[50-52]. Interestingly, Liu et al. delineated a rapid glucoregulatory circuit: paraventricular nucleus (PVN) corticotropin-releasing hormone (CRH) neurons → ventromedial nucleus of the hypothalamus (VMH) → raphe pallidus → sympathetic nerves → liver (the hypothalamic-sympathetic-liver axis), distinct from the classic HPA axis that raises blood sugar comparatively slowly[53]. In adrenalectomized mice, physical stressors induced rapid hyperglycemia and increased activity of key gluconeogenic enzymes in the liver, indicating that HSL axis activation promotes glucose release via hepatic gluconeogenesis[54]. On the other hand, the PNS generally promotes anabolic processes, enhancing glucose storage and insulin sensitivity[55,56]. Acute hepatic vagotomy or central vagal stimulation in rats significantly increased arterial and portal venous insulin levels or inhibited insulin secretion, respectively, suggesting a role for hepatic vagal nerves in insulin secretion regulation[57]. Complete hepatic denervation may abolish net hepatic glucose uptake[58], as observed in liver transplant recipients during hypoglycemia[59].
In lipid metabolism, the SNS and PNS collectively maintain hepatic lipid homeostasis[60]. The SNS primarily promotes hepatic lipid accumulation. Elevated hepatic sympathetic tonus drives steatosis by simultaneously accelerating free fatty acid uptake and intrahepatic de novo lipogenesis[61]. Consistent with this, sympathetic denervation has been shown to reverse established hepatic steatosis in obese mice[61] and to reduce VLDL-TG secretion in dyslipidemic rats[62]. Together, these findings indicate that chronic sympathetic overactivity is a central pathogenic factor in the development of MASLD[63-65]. In contrast, the PNS mediates anti-steatotic signals from the brain. Adipose tissue-derived leptin translocates across the blood-brain barrier to exert central actions on the dorsal vagal complex, subsequently enhancing triglyceride export and suppress hepatic de novo lipogenesis through vagal efferent signaling[66]. These centrally mediated leptin effects depend on intact hepatic vagal innervation and are absent in liver-transplanted recipients lacking neural input to the graft[67]. Vagal sensory neurons innervating the liver are required for the development of diet-induced hepatic steatosis[68].
The ANS precisely regulates bile acid synthesis and secretion through bidirectional actions of sympathetic and parasympathetic nerves, serving as a crucial component in maintaining bile acid pool stability. The PNS enhances cholecystokinin-induced Cl-/HCO3- exchange in cholangiocytes by releasing acetylcholine, synergizing with secretin to promote bile secretion[69]. The SNS primarily acts on cholangiocyte α1-adrenergic receptors by virtue of norepinephrine, reducing bile secretion[70]. This dual innervation ensures bile secretion adapts to varying physiological demands. Animal studies in Wistar rats demonstrate that vagus nerve transection significantly elevates levels of bile acids such as TMDCA, GHDCA, ω-MCA, α-MCA, CA, isoCDCA, CDCA, DCA, 7-oxo-HDCA, and 12-oxo-CA. This mechanism involves the vagus nerve activating the intestinal FXR-Fgf15 signaling pathway while simultaneously suppressing hepatic expression of the bile acid synthase Cyp7a1[71]. Recent studies further confirm that parasympathetic nerve terminals in the human hepatic biliary system release vesicular acetylcholine transporter (VAChT, a parasympathetic marker)[72], providing direct evidence for cholinergic nerves directly regulating bile secretion.
Although the ENS is anatomically localized within the gastrointestinal tract, it exerts indirect regulatory effects on liver function via a complex, multi-component gut-liver axis. Central to this regulation are EECs, which act as specialized chemosensory epithelial cells capable of detecting luminal chemical and microbial stimuli. Upon activation, EECs secrete neuroactive hormones, including serotonin, glucagon-like peptide-1 (GLP-1), and CCK, that signal to the central nervous system primarily by means of vagal afferent fibers[68]. Subsequent central integration modulates autonomic outflow to the liver and affects hepatic metabolic, synthetic, and detoxification functions[73]. Concurrently, the gut microbiota contributes to this regulatory network by producing bioactive metabolites that modulate EEC activity, maintain intestinal barrier integrity, and shape both local and systemic immune responses, collectively impacting hepatic physiology and pathophysiology[74,75]. Collectively, current evidence indicates that ENS-mediated regulation of liver function involves bidirectional crosstalk among neural, endocrine, microbial, and immune components. However, the precise molecular and neural circuit mechanisms underlying this inter-organ communication remain incompletely understood.
ANS IN THE GUT-LIVER METABOLIC AXIS
Extensive clinical studies indicate that 60%-80% of patients with PSC have IBD, while 2%-14% of IBD patients are diagnosed with PSC[76]. Whether IBD is an extraintestinal manifestation of PSC or a closely related comorbidity remains controversial. However, evidence strongly supports that gut-liver communication is involved in shared metabolic and inflammatory pathways, though the precise underlying mechanisms are still unclear[77,78]. This article summarizes how ANS enables interorgan dialogue among the intestine, liver and brain through immunity, metabolism and intestinal microbes, with particular focus on metabolic regulation [Figure 3].
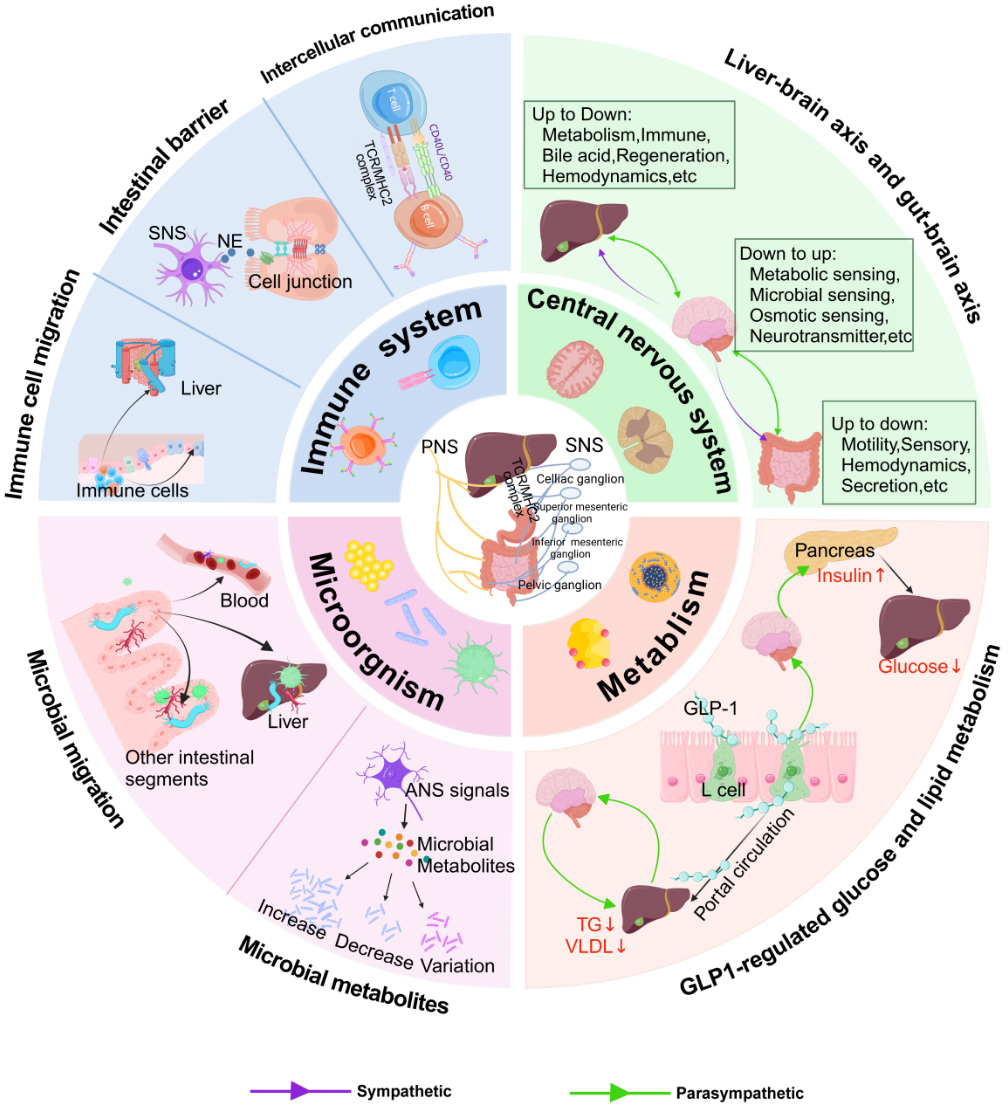
Figure 3. The ANS orchestrates gut-liver metabolic homeostasis through integrated neuro-metabolic pathways. The ANS modulates gut-liver metabolic communication via four interconnected routes: glucose and lipid metabolism, immunometabolism, microbial metabolism, and CNS metabolic integration. In metabolic regulation, the ANS controls hepatic and intestinal glucose/lipid metabolism via GLP-1 signaling and direct neural inputs, regulating gluconeogenesis, lipogenesis, and energy balance. In immunometabolism, the ANS shapes the metabolic microenvironment of gut and liver by modulating immune cell trafficking and barrier integrity, influencing insulin sensitivity and nutrient processing. Regarding microbial metabolism, autonomic innervation affects microbial metabolite production (SCFAs, bile acids), which subsequently regulate host metabolism, insulin secretion, and energy expenditure. Parasympathetic pathways convey ascending signals to the central nervous system, which integrates this information and coordinates sympathetic and parasympathetic efferent outputs to target organs. These neural circuits collectively regulate gut-liver metabolic homeostasis. Image created with cnsknowall.com. SNS: Sympathetic nervous system; ANS: autonomic nervous system; NE: norepinephrine; TG: triglyceride; VLDL: very low density lipoprotein; GLP-1: glucagon-like peptide-1; TCR: T cell receptor; MHC II: major histocompatibility complex class II.
Autonomic regulation of gut-liver axis immunometabolic homeostasis
Sympathetic and parasympathetic nerves jointly constitute a refined neuro-immune dialogue network within the gut-liver axis by regulating barrier function, immune cell migration, and intercellular communication, all of which have secondary effects on tissue metabolic function.
Maintaining intestinal barrier integrity relies on stable expression of epithelial tight junction proteins[79]. Sympathetic-derived norepinephrine engages β2-ARs on intestinal epithelial and immune cells populations, subsequently driving the downregulation of tight junction proteins, compromising epithelial barrier integrity, and facilitates the translocation of microbial products into the circulation[80]. At the cellular level, sympathetic-derived NE targets dendritic cell populations via β-AR pathways, a signaling axis that critically patterns subsequent T cell activation and dictates the localized immunometabolic microenvironment[81]. Sympathetic signaling additionally regulates immune cell migration by altering portal vein blood flow and metabolic substrate delivery. Sympathetic activation induces intestinal vasoconstriction, reduces immune cell infiltration into the gut and suppresses local immune responses[82]. Clinical studies have identified memory T cells of common origin in intestinal and hepatic samples from PSC-IBD patients, suggesting bidirectional migration of immune cells between the gut and liver that jointly shapes the immune and metabolic microenvironments of both organs[83].
In contrast to the PNS, the vagus nerve exerts protective effects in the gut-liver axis by the “cholinergic anti-inflammatory pathway.” Within the intestine, the vagus nerve can sense antigens through high-affinity Immunoglobulin E receptors and release glutamate to modulate dendritic cell function, suppressing Th2-skewed inflammation[84]. Following injury, enteric glial cells are critical for transmitting vagal anti-inflammatory signals, enhancing barrier repair and limiting immune cell recruitment[85]. Concurrently, gut-derived microbial metabolites and luminal inflammatory signals are monitored by hepatic vagal sensory afferents. Through the liver-brain-gut neural arc, these signals are transmitted to the solitary nucleus in the brainstem. Subsequently, through the vagus nerve’s parasympathetic efferent pathway, the vagus nerve modulates the abundance of gut immune cells, forming an indirect pathway influencing the hepatic immune and metabolic environment[86].
Autonomic regulation of gut-liver axis metabolic homeostasis
Traditionally, the liver was regarded as a passive metabolic executor, and the gut was viewed primarily as an absorptive organ. Recent paradigms have illuminated the liver not merely as a passive metabolic sink, but as an active sensory organ participating in neuroendocrine feedback loops, operating in concert with extensive gut-CNS conduits via EECs, the ENS, and the microbiota[87-90]. This gut-liver-brain triad forms the physiological basis for metabolic homeostasis, whose disruption precipitates various metabolic disorders such as MASLD[91,92].
A key mediator in this circuit is the gut hormone GLP-1, whose actions illustrate the integrated neural control of metabolism across organs. Upon intraluminal nutrient engagement, mucosal L-cells liberate GLP-1, which subsequently docks with cognate receptors localized on vagal afferent terminals within the intestinal wall and portal vein, initiating a neural signal to the brainstem[93]. The metabolic actions of GLP-1 receptor agonists, including the inhibition of gastric emptying and stimulation of insulin secretion, are mediated through the vagus nerve[94]. Evidence shows that knockdown of GLP-1 receptors in vagal afferent neurons blunts these effects, while vagal nerve stimulation can mimic them by enhancing vagal activity and increasing GLP-1 release[95-97]. The same neural circuit directly controls liver lipid metabolism[98]. In another key experiment, researchers injected GLP-1 into the portal vein and observed a significant reduction in blood triglycerides and hepatic VLDL secretion in both hamsters and mice. When the vagal afferent pathway was interrupted either surgically or pharmacologically, this lipid-lowering effect disappeared[98]. Moreover, the anti-lipemic effect of portal GLP-1 requires not only intact vagal signaling but also efferent changes in sympathetic tone[98]. This indicates that the brain’s processing of vagal GLP-1 input includes adjustments to SNS outflow. Regarding bile acid metabolism, activation of intestinal FXR transmits signals to the liver by virtue of vagus nerve and regulates cholesterol 7α-hydroxylase expression, forming a neural component of the enterohepatic feedback loop[99]. Recently, single-cell technologies have advanced the field. Newly identified vagal CART⁺ neuron subsets regulate insulin and hepatic gluconeogenesis, a mechanism linked to commensal microbiota dysbiosis or depletion, representing a breakthrough in understanding the microbiota-gut-liver axis[100].
Taken together, these paradigms establish the ANS as a critical rheostat that fine-tunes gut-liver metabolic homeostasis utilizing an integrated neuroendocrine framework.
Autonomic regulation of the gut-liver axis via microbial metabolism
As a core component of the gut-liver axis, the gut microbiota function as a critical determinant in the dynamic balance between the intestine and liver[88,101]. The ANS, in turn, reshapes the microbiota-gut-liver network via direct neural modulation, metabolite intervention, and regulation of microbial translocation, emerging as a novel therapeutic target for metabolic diseases.
Microbial metabolites
Gut metabolites (e.g., SCFAs, secondary bile acids, tryptophan derivatives, neurotransmitters) are small molecules produced by the microbiota that serve as key metabolic mediators[102]. Although of microbial origin, these metabolites reciprocally shape microbial ecology by the means of microenvironmental modifications, gene expression modulation, quorum sensing disruption, and metabolic reprogramming. The ANS can influence these microbial metabolites, regulate microbial composition and function, and affect host metabolism. For example, the ANS can modulate microbial bile acid metabolism by activating receptors like FXR and TGR5. Employing PNS signaling, Lactiplantibacillus plantarum increases tauroursodeoxycholic acid (TUDCA) levels and reduces lithocholic acid (LCA) levels, modulating microbial metabolic pathways and promoting beneficial bacteria (e.g., lactobacilli) growth[103]. SNS-derived NE, via the Adra2a signaling axis, lowers cAMP to inhibit calcium flux, suppressing GLP-1 release and raising blood glucose[104]. Knockout of Adra2a or treatment with the GLP-1R agonist exendin-4 lowers blood glucose in mice, affecting microbial carbon sources[104]. The vagus nerve promotes primary bile acid secretion by activating hepatic FFAR3. These bile acids stimulate microbial conversion to secondary bile acids in the gut, forming a feedback loop regulating hepatic lipid metabolism[105]. The ANS can shape the gut microbial ecosystem by influencing the production and profile of key metabolites.
Microbial translocation
The ANS critically governs the gut-liver axis by modulating microbial translocation and its attendant metabolic sequelae, a process sustained by the tripartite regulation of epithelial barrier integrity, immune microenvironment dynamics, and microbial ecology. Long-term chronic stress disrupts the intestinal barrier via hyperactivation of SNS signaling, thereby precipitating the translocation of bacterial endotoxins and opportunistic pathogens[106]. However, SNS signaling also participates in maintaining intestinal mucosal immune homeostasis, preserving epithelial barrier integrity, and supporting antimicrobial defense. This functional duality is highly context-dependent, tailored by the precise nature of the physiological or pathological stimulus (e.g., acute or chronic, the magnitude and duration of stimulation). Pharmacological depletion of sympathetic innervation in the lamina propria and mucosa increased inflammatory cell infiltration (CD68 + CD86 macrophages, Ly6C+ monocytes) and promoted the expression of pro-inflammatory cytokines (IL-1β, TNF-α, IFN-γ) in the gut. The translocation of intestinal bacteria into the liver via a compromised epithelial barrier represents a seminal and pervasive event in the pathogenesis of MASLD[107]. Studies in MASLD mouse models demonstrated that Escherichia coli (E. coli) activates the TLR5/MYD88/TWIST1 pathway in liver sinusoidal endothelial cells through flagellin signaling[107]. These changes promote steatosis, inflammation, and fibrosis in MASLD, highlighting the link between microbial translocation and metabolic dysfunction[107]. Clinical studies have shown that reduced vagus nerve tone in patients with cirrhosis is significantly associated with intestinal flora imbalance and increased risk of hepatic encephalopathy, while vagus nerve stimulation (VNS) can improve liver injury by restoring the intestinal barrier and preserving metabolic homeostasis[108-111]. These findings reveal the potential therapeutic value of ANS in gut-liver axis disease by rebalancing microbial translocation homeostasis and protecting metabolic function.
Autonomic regulation of the gut-liver axis via the CNS
The gut-brain axis is a complex bidirectional network that links brain regions involved in emotion and cognition with peripheral gut function and coordinates metabolic homeostasis[5,112]. The gut signals up to the brain through spinal and vagal visceral afferents and receives “top-down” signals under the mediation of sympathetic and parasympathetic efferents, forming a circuit for metabolic regulation[113]. Vagal afferents from the gut smooth muscle transmit information regarding nutrient status to the CNS[114,115]. Clinically, vagotomy performed for peptic ulcer disease has been associated with an increased incidence of mental health disorders. In animal studies, transection of subdiaphragmatic vagal afferents induces anxiety- and fear-related behaviors[116,117], suggesting that disruption of gut-derived vagal signaling can trigger psychiatric symptoms. Diverging from the bidirectional modality of the vagus nerve, the SNS operates predominantly as an obligate efferent axis, dictating gastrointestinal motility and glandular output via norepinephrine release during stress states.
Similarly, within the liver-brain axis, hepatic vagal afferents indirectly sense the hepatic microenvironment and relay this information to the NTS, which subsequently modulates hepatic parasympathetic outflow[5,116,118]. Exogenous vagal reflex activity can connect hepatic vagal afferents, the brainstem, vagal efferents, and enteric neurons[118]. Hepatic ischemia-reperfusion (HIR) injury often induces systemic inflammation, depression-like behavior, and reduced prefrontal synaptic protein levels in mice[102]. Notably, subdiaphragmatic vagotomy attenuates these symptoms[119], implicating the vagal signaling in this pathological process. As in the gut-brain axis, parasympathetic pathways mediate bidirectional communication in the liver-brain axis, while sympathetic signaling predominantly provides efferent output[120]. Chronic stress further drives CRH+ CeM → PVN projections, sustaining hepatic sympathetic outflow. This process compromises intrahepatic sympathetic fibers, elevated circulating norepinephrine levels, β3-adrenergic receptor downregulation, and impaired cAMP signaling in hepatocytes, ultimately contributing to hepatic catecholamine resistance and MASLD progression[10].
Interactions among the brain, liver, and gut involve highly complex regulatory mechanisms[102]. Currently, the liver-brain-gut neural reflex arc proposed by Teratani et al. has gained wide recognition[102]. Specifically, hepatic vagal afferents sense the gut microenvironment and signal to the NTS[102]. Central integration within the NTS subsequently modulates peripheral vagal and enteric outputs, culminating in the induction and phenotypic preservation of intestinal pTregs[86]. This reflex arc provides a representative example of how CNS-processed autonomic signaling coordinately regulates immune homeostasis along the gut-liver axis.
TARGETING THE ANS FOR INTERVENTION OF GASTROINTESTINAL DISEASE
Emerging ANS-targeted therapies primarily involve VNS, which can be categorized into invasive and noninvasive approaches. Invasive VNS requires surgical implantation of electrodes around the cervical vagus nerve, whereas noninvasive approaches, especially transcutaneous auricular VNS (taVNS), have attracted increasing research interest because of their safety and convenience. Other noninvasive techniques include transcutaneous cervical VNS and percutaneous electrical nerve field stimulation[121].
In IBD, taVNS demonstrates clear anti-inflammatory effects. In rodent colitis models, it reduces disease activity index, improves colonic injury, and lowers pro-inflammatory cytokine levels[122,123]. Small clinical studies show that taVNS can induce clinical remission in some patients with mild-to-moderate Crohn’s disease (CD) or ulcerative colitis (UC), accompanied by decreased fecal calprotectin[124]. Part of the vagus nerve’s anti-inflammatory action is mediated by the “splenic-vagal pathway,” as electrical stimulation of the splenic nerve bundle also ameliorates colitis in mice[125]. In gastrointestinal motility disorders, taVNS shows significant prokinetic effects that enhance nutrient processing and metabolism. In functional dyspepsia (FD) patients, it increases gastric volume and restores gastric slow-wave rhythm[126]. In constipation-predominant irritable bowel syndrome (IBS-C), it increases bowel movement frequency, alleviates abdominal pain, and improves anorectal sensation and mood[127]. These benefits are attributed to enhanced vagal tone and restored parasympathetic regulation of gastrointestinal motility after taVNS. Experimental central vagal activation can prevent postoperative gastroparesis[128] [Table 2]. In liver diseases, noninvasive VNS improves metabolic markers in patients with metabolic syndrome and has demonstrated potential efficacy in reducing hepatic steatosis, inflammation, and fibrosis [Table 3][129,130].
ANS interventions in gut diseases
| Author | Intervention | Disease | Study design | Participants | Location | Major results |
| Zhu et al.[126] | taVNS | FD | double-blind, randomized, sham-controlled, small pilot RCT | 18:18 | bilateral auricular cymba concha areas | Engagement of vagal pathways was associated with improved gastric accommodation and stabilization of gastric pacemaker activity, supporting therapeutic utility in nonservice FD |
| Shi et al.[138] | taVNS | FD | randomized, sham-controlled, RCT | 10-Hz tins group:25-Hz tens group: sham group = 101:99:100 | tragus of the left ear | A higher proportion of responders and adequate symptom relief was observed in active stimulation groups relative to sham after 4 weeks |
| Shi et al.[127] | taVNS | IBS-C | randomized, sham-controlled, small pilot RCT | 21:19 | bilateral auricular cymba concha areas | Concurrent improvement in bowel function and abdominal pain, consistent with modulation of gut-brain signaling |
| Kovacic et al.[139] | PENFS | Abdominal pain-related functional gastrointestinal disorders in adolescents | double-blind, randomized, sham-controlled, RCT | 51:47 | dorsal and ventral aspects of the ear within 1-1·5 mm of the vascular branches, but not on main arterial branches | Sustained reduction in abdominal pain-related symptom burden in adolescents with functional gastrointestinal disorders |
| Krasaelap et al.[140] | PENFS | IBS in adolescents | double-blind, randomized, sham-controlled, small pilot RCT | 27:23 | vascular branches of the outer ears | Reduction in pain severity accompanied by improved patient-reported well-being in adolescent IBS |
| Sahn et al.[124] | taVNS | IBD (children) | single-blind, randomized, sham-controlled, small pilot RCT | 11:11 (CD:UC=10:12) | cymba conchae of the external left ear | Favorable safety profile with concurrent symptom improvement and decreased fecal calprotectin, indicating reduced intestinal inflammatory activity |
| Huang et al.[141] | TEA | IBS-C | single-blind, randomized, sham-controlled, small pilot RCT | 26:26 | bilateral PC6 and ST36 | Acceleration of colonic transit and attenuation of rectal hypersensitivity, consistent with autonomic regulatory involvement |
| Zhou et al.[142] | TEA | CC | single-blind, randomized, sham-controlled, small pilot RCT | 22:22 | ST36 | Sustained alleviation of constipation linked to enhanced vagal tone and normalization of rectal sensory function |
| Liu et al.[143] | EA | CSFC | single-blind, randomized, parallel, sham-controlled, RCT | 536:539 | bilateral acupoints of ST25, SP14, andST37 | Increased frequency of complete spontaneous bowel movements with a favorable safety profile following 8-week EA |
| Liu et al.[144] | EA | SCC | multicenter, randomized, controlled, noninferiority, RCT | 280:280 | bilateral ST25, SP14 and ST37 | Clinical efficacy comparable to prucalopride with prolonged post-treatment benefit and good tolerability |
| Iqbal et al.[145] | tSNS | CFC | open-label, exploratory research | 16 | the entire sacrum | No meaningful short-term improvement in chronic constipation |
| Iqbal et al.[146] | tSNS | CFC | open-label, exploratory research | 18 | behind the medial malleolus where the tibial nerve is most superficial (bilateral) | Clinical benefit observed in a subset of patients, particularly those with less severe disease phenotypes |
| Gokce et al.[147] | tTNS | CFC (geriatric patients) | open-label, exploratory research | 44 | TENS electrodes were placed below the medial malleolus at approximately 15-cm above the medial malleolus (bilateral) | Significant improvements in defecatory dysfunction, colonic inertia, and pain, with sustained analgesic effects |
| Chase et al.[148] | IFT | chronic treatment-resistant constipation and soiling (children) | open-label, exploratory research | 8 | Abdominal/paraspinal(T9 and L2) | Marked reduction in soiling and increased spontaneous bowel activity, with partial durability of response |
| Clarke et al.[149] | IFT | STC | randomized, sham-controlled, small pilot RCT | 22:8 | Abdominal/paraspinal(T9 and L2) | Enhanced colonic transit in pediatric slow transit constipation. |
| Clarke et al.[150] | IFT | STC | double-blind, randomized, sham-controlled, small pilot RCT | 16:17 | Abdominal/paraspinal(T9 and L2) | Improvements in child-reported quality of life and functional scores restricted to active treatment |
| Ismail et al.[151] | TES | STC (children) | open label, exploratory research | 11 | Abdominal/paraspinal(T9 and L2) | Home-based TES associated with improved bowel function in treatment-resistant cases |
| Yik et al.[152] | TES | STC (children) | open-label, exploratory research | 29 | Abdominal/paraspinal(T9 and L2) | Increased defecation frequency and reduced soiling, with approximately half of patients responding |
| Queralto et al.[153] | IFT | STC | prospective, exploratory research | 11 | Abdominal/paraspinal(T9 and L2) | Preliminary evidence supporting IFT as a noninvasive option for STC |
| Clarke et al.[154] | IFC | STC (children) | open-label, exploratory research | 8 | Abdominal/paraspinal(T9 and L2) | Increased propagated colonic motor activity with effects persisting for months |
| YYang et al.[155] | TES | STC (female) | assessor-blinded, randomized, controlled, small pilot RCT | 14:14 | Abdominal/paraspinal(T9 and L2) | Symptom reduction in female STC patients |
| Moore[156] | IFT | STC (female) | single-blind, randomized, sham-controlled, small pilot RCT | 17:16 | NR | Sustained reduction in constipation severity with improved quality of life |
| Xiao et al.[157] | TENS | IBS-D, IBS-C, FC | open-label, exploratory research | IBS-D:IBS-C:FC:HC=24:20:30:30 | LI4,ST36, UB57 | Clinical efficacy observed in IBS-D. |
| Xiao et al.[158] | TEA | FC | randomized, patient-blinded, sham-controlled, small pilot RCT | 27:269:9 | ST36 | Improved bowel function associated with enhanced rectal sensation and parasympathetic activity |
| Coban et al.[159] | IFC | IBS | double-blind, randomized, sham-controlled, small pilot RCT | 29:29 | Abdominal/paraspinal(T9 and L2) | Significant improvement in IBS symptoms and quality of life |
ANS interventions in liver diseases
| Author | Intervention | Disease | Study design | Participants | Location | Major results |
| Adams et al.[160] | Liver transplantation | FAP | follow-up study | 45 | NA | Reduction in circulating mutant transthyretin and slowed axonal degeneration, with limited impact on autonomic dysfunction and persistence of systemic symptoms |
| Zhao et al.[161] | EA | MASLD | randomized, patient-blinded, sham-controlled, RCT | 72:72 | CV12, CV4, ST25, SP15, LR13, LI4, ST36, SP6, LR3 | ChiCTR2200060353 |
| Fu et al.[162] | EA | MASLD | randomized, sham-controlled, small pilot RCT | 49:49 | RN12, RN9, CV4, RN6, ST24, ST25, SP15, GB26, ST40, ST36, SP6 | ChiCTR2300075701 |
| Metz et al.[67] | Metreleptin | MASLD | randomized, placebo-controlled, small pilot RCT | 13:9 | NA | Anti-steatotic effects mediated via central-vagal-hepatic signaling, independent of caloric intake |
| Jakovljevic et al.[163] | Resistance exercise | MASLD | randomized, controlled, small pilot RCT | 8:9 | NA | Improved autonomic balance and hemodynamic responses during submaximal exercise |
Beyond neural stimulation itself, combined microbiota-neural interventions show synergistic potential for metabolic improvement. The gut microbiota, via metabolites like SCFAs, interacts with the immune system and profoundly influences vagal activity, forming a bidirectional “microbiota-gut-brain axis” that regulates host metabolism. For instance, heat-inactivated Pediococcus acidilactici R037 enhances sympathetic activity in white adipose tissue, promoting lipolysis and reducing serum TG, suggesting utility in primary prevention of coronary artery isease and other metabolic conditions[132]. In alcoholic and non-alcoholic fatty liver diseases, reduced vagal tone correlates with gut dysbiosis, impaired barrier function, and liver inflammation, suggesting that restoring parasympathetic balance could be a novel therapeutic avenue for these metabolic liver diseases[133-135]. Future strategies are shifting towards closed-loop bioelectronic medicine systems that dynamically adjust stimulation parameters based on real-time physiological markers, such as heart rate variability, electrogastrogram, and inflammatory biomarkers, thereby enabling more precise and adaptive neuromodulation.
Challenges and future perspectives
Despite the therapeutic promise, neuromodulation strategies for gut-liver diseases face challenges. A core technical bottleneck is a lack of stimulation specificity. Current studies vary considerably in stimulation frequency, target sites, and treatment duration, highlighting the need for large-scale randomized controlled trials to establish universal protocols[121]. In addition, the metabolic effects of sympathetic activation are highly context dependent. In the tumor microenvironment, SNS-derived NE can promote colorectal cancer progression via a β2-ARs-mediated positive feedback loop that stimulates cancer-associated fibroblasts to secrete nerve growth factor, potentially altering tumor metabolism[136]. These findings emphasize the importance of precise neural targeting and a clearer understanding of the diverse roles of sympathetic signaling in metabolic regulation. This issue is pronounced in the emerging field of “microbiota-neural combination therapy.” It remains unclear how neural stimulation precisely affects specific gut microbes and how microbial metabolites feedback onto neural pathways. This knowledge gap hinders the rational design of combined therapies.
Future progress requires integrated multi-organ monitoring systems capable of simultaneously assessing neural activity, gut motility, microbiome, and inflammatory status. In parallel, advances in highly selective neuromodulation technologies are needed to precisely target functional neural subpopulations. Recent studies have shown that distinct sympathetic neuron subsets separately regulate gastrointestinal motility and secretion[137], providing opportunities for more targeted interventions while minimizing off-target effects. Importantly, substantial interspecies differences must also be considered. The distribution of hepatic autonomic innervation differs between rodents and humans, limiting the direct translational applicability of rodent-based findings[44]. Collectively, improving personalized precision medicine approaches is a promising direction in the future. Greater integration of neuroscience, immunology, microbiology, and clinical medicine is essential for developing safe and effective targeted therapies for gut-liver disorders.
CONCLUSION
The ANS functions as a central regulator of the gut-liver axis, integrating metabolic, immune, and microbial homeostasis through its sympathetic, parasympathetic, and enteric branches[5]. Dysregulation of this neural control likely contributes to the shared pathogenesis and frequent co-occurrence of intestinal and hepatic diseases, particularly those involving metabolic dysfunction. Targeting the ANS, especially through vagus nerve modulation such as taVNS, has emerged as a promising therapeutic strategy. However, major challenges exist in stimulation specificity and precise regulation. Further investigation is required to clarify how neural circuits coordinate immune and metabolic signaling across the gut-liver axis. Ultimately, realizing the therapeutic potential of ANS-targeted intervention requires precision medicine approaches targeting the vagus nerve, tailored to complex gut-liver disorders.
DECLARATIONS
Authors’ contributions
Writing-original draft, Writing-review & editing: Sun R, He JL, Cheng Y
Writing-review & editing, Project administration: Yu SR, Zhang MN, Yang M, Chen K
Funding acquisition, Conceptualization, Supervision, Writing-review & editing: Wu MY, Chai J
Availability of data and materials
Not applicable.
AI and AI-assisted tools statement
Not applicable.
Financial support and sponsorship
This work was supported by Noncommunicable Chronic Diseases-National Science and Technology Major Project (2026ZD0557000), Prevention and Control of Emerging and Major Infectious Diseases-National Science and Technology Major Project (2025ZD101906400) awarded to Chai J, the National Natural Science Foundation of China (No. 82570651), Outstanding Youth Talent Program of Southwest Hospital (2025YQBJ-5) awarded to Wu MY.
Conflicts of interest
All authors declared that there are no conflicts of interest.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Copyright
© The Author(s) 2026.
REFERENCES
1. Devarbhavi H, Asrani SK, Arab JP, Nartey YA, Pose E, Kamath PS. Global burden of liver disease: 2023 update. J Hepatol. 2023;79:516-37.
2. Griffin C, Agbim U, Ramani A, Shankar N, Kanwal F, Asrani SK. Underestimation of cirrhosis-related mortality in the medicare eligible population, 1999-2018. Clin Gastroenterol Hepatol. 2023;21:223-225.e3.
3. Li Q, Kou Y, Zhang M, et al. Sex-specific and socioeconomic disparities in the global burden of inflammatory bowel disease in 204 countries, 1990-2021: projections to 2050. Front. Immunol. 2026;16:1673212.
4. Ali B, Samuel R, Kramer JR, et al. Evolving burden of metabolic dysfunction-associated steatotic liver disease and its complications in a US nationwide healthcare system. Hepatol Commun. 2026;10.
5. Gu Y, Wu C. The gut-liver axis modulates intestinal immune homeostasis. Mucosal Immunol. 2026;19:1470-80.
6. Devkota S, Wang Y, Musch MW, et al. Dietary-fat-induced taurocholic acid promotes pathobiont expansion and colitis in Il10-/- mice. Nature. 2012;487:104-8.
7. Yan J, Zeng Y, Guan Z, et al. Inherent preference for polyunsaturated fatty acids instigates ferroptosis of Treg cells that aggravates high-fat-diet-related colitis. Cell Reports. 2024;43:114636.
8. Rodriguez-duque JC, Calleja JL, Iruzubieta P, et al. Increased risk of MAFLD and liver fibrosis in inflammatory bowel disease independent of classic metabolic risk factors. Clin Gastroenterol Hepatol. 2023;21:406-414.e7.
9. Sourianarayanane A, Garg G, Smith TH, Butt MI, Mccullough AJ, Shen B. Risk factors of non-alcoholic fatty liver disease in patients with inflammatory bowel disease. J Crohns Colitis. 2013;7:e279-85.
10. Wu S, Liu J, Huang S, Guo Y, Bi Y. Chronic stress induces hepatic steatosis via brain-hepatic sympathetic axis mediated catecholamine resistance. Int. J. Biol. Sci. 2026;22:1407-24.
12. Thayer JF, Lane RD. The role of vagal function in the risk for cardiovascular disease and mortality. Biol Psychol. 2007;74:224-42.
13. Wang T, Tufenkjian A, Ajijola OA, Oka Y. Molecular and functional diversity of the autonomic nervous system. Nat Rev Neurosci. 2025;26:607-22.
14. Mawdsley JE, Rampton DS. Psychological stress in IBD: new insights into pathogenic and therapeutic implications. Gut. 2025;54:1481-91.
15. Luo Q, Liu P, Dong Y, Qin T. The role of the hepatic autonomic nervous system. Clin Mol Hepatol. 2023;29:1052-5.
16. Albillos A, De Gottardi A, Rescigno M. The gut-liver axis in liver disease: Pathophysiological basis for therapy. J Hepatol. 2020;72:558-77.
17. Dinan TG, Cryan JF. Regulation of the stress response by the gut microbiota: Implications for psychoneuroendocrinology. Psychoneuroendocrinology. 2012;37:1369-78.
18. Sgambato D, Miranda A, Ranaldo R, Federico A, Romano M. The role of stress in inflammatory bowel diseases. Curr Pharm Des. 2017;23.
19. Zhai S, Zhu Y, Wang X, et al. Development of a predictive model for radiation pneumonitis based on plasma exosomal miR-200b-5p. Front. Oncol. 2025;15:1516348.
20. Basics of autonomic nervous system function. Clinical Neurophysiology: basis and technical aspects. Elsevier; 2019. pp. 407-18.
22. Sanders KM, Mutafova-Yambolieva VN. Neurotransmitters responsible for purinergic motor neurotransmission and regulation of GI motility. Auton Neurosci. 2021;234:102829.
24. Janig W, Mclachlan EM. Organization of lumbar spinal outflow to distal colon and pelvic organs. Physiol Rev. 1987;67:1332-404.
25. Simmons M. The complexity and diversity of synaptic transmission in the prevertebral sympathetic ganglia. Prog Neurobiol. 1985;24:43-93.
26. Altschuler SM, Escardo J, Lynn RB, Miselis RR. The central organization of the vagus nerve innervating the colon of the rat. Gastroenterology. 1993;104:502-9.
27. De Groat WC, Nadelhaft I, Milne RJ, Booth AM, Morgan C, Thor K. Organization of the sacral parasympathetic reflex pathways to the urinary bladder and large intestine. J Auton Nerv Syst. 1981;3:135-60.
28. Vizzard MA, Brisson M, De Groat WC. Transneuronal labeling of neurons in the adult rat central nervous system following inoculation of pseudorabies virus into the colon. Cell Tissue Res. 2000;299:9-26.
29. Shields RW. Functional anatomy of the autonomic nervous system. J Clin Neurophysiol. 1993;10:2-13.
30. Zhu YF, Wang X, Lowie B, et al. Enteric sensory neurons communicate with interstitial cells of Cajal to affect pacemaker activity in the small intestine. Pflugers Arch - Eur J Physiol. 2013;466:1467-75.
31. Ellis M, Chambers JD, Gwynne RM, Bornstein JC. Serotonin and cholecystokinin mediate nutrient-induced segmentation in guinea pig small intestine. Am J Physiol Gastrointest Liver Physiol. 2013;304:G749-61.
32. Travagli RA, Hermann GE, Browning KN, Rogers RC. Brainstem circuits regulating gastric function. Annu Rev Physiol. 2006;68:279-305.
33. Spencer NJ, Dinning PG, Brookes SJ, Costa M. Insights into the mechanisms underlying colonic motor patterns. J Physiol. 2016;594:4099-116.
34. Furness JB, Callaghan BP, Rivera LR, Cho H. the enteric nervous system and gastrointestinal innervation: integrated local and central control. In: Lyte M, Cryan JF, Editors. Microbial Endocrinology: The Microbiota-Gut-Brain Axis in Health and Disease. New York, NY: Springer New York; 2014. pp. 39-71.
35. Parker DR, Wiklendt L, Humenick A, et al. Sympathetic pathways target cholinergic neurons in the human colonic myenteric plexus. Front Neurosci. 2022;16:863662.
37. Horiuchi J, Mcdowall L, Dampney R. Differential control of cardiac and sympathetic vasomotor activity from the dorsomedial hypothalamus. Clin Exp Pharmacol Physiol. 2006;33:1265-8.
38. Hao S, Dulake M, Espero E, Sternini C, Raybould HE, Rinaman L. Central Fos expression and conditioned flavor avoidance in rats following intragastric administration of bitter taste receptor ligands. Am J Physiol Regul Integr Comp Physiol. 2009;296:R528-36.
39. Sartor DM, Verberne AJM. Cholecystokinin selectively affects presympathetic vasomotor neurons and sympathetic vasomotor outflow. Am J Physiol Regul Integr Comp Physiol. 2002;282:R1174-84.
40. Holmes GM, Tong M, Travagli RA. Effects of brain stem cholecystokinin-8s on gastric tone and esophageal-gastric reflex. Am J Physiol Gastrointest Liver Physiol. 2009;296:G621-31.
41. Effects of gut bacteria and their metabolites on gut health of animals. Elsevier; 2024. pp. 223-52.
42. Agibalova T, Hempel A, Maurer HC, et al. Vasoactive intestinal peptide promotes secretory differentiation and mitigates radiation-induced intestinal injury. Stem Cell Res Ther. 2024;15:348.
43. Sun M, Wan Y, Shi M, Meng Z, Zeng W. Neural innervation in adipose tissue, gut, pancreas, and liver. Life Metabolism. 2023;2:load022.
44. Miller BM, Oderberg IM, Goessling W. Hepatic nervous system in development, regeneration, and disease. Hepatology. 2021;74:3513-22.
45. Zou J, Li J, Wang X, Tang D, Chen R. Neuroimmune modulation in liver pathophysiology. J Neuroinflammation. 2024;21:188.
46. Luo Q, Liu P, Dong Y, Qin T. The role of the hepatic autonomic nervous system. Clin Mol Hepatol. 2023;29:1052-5.
48. Carty JRE, Devarakonda K, O’connor RM, et al. Amygdala-liver signalling orchestrates glycaemic responses to stress. Nature. 2025;646:697-706.
49. Shimazu T. Innervation of the liver and glucoregulation: Roles of the hypothalamus and autonomic nerves. Nutrition. 1996;12:65-6.
50. Abot A, Lucas A, Bautzova T, et al. Galanin enhances systemic glucose metabolism through enteric Nitric Oxide Synthase-expressed neurons. Mol Metab. 2018;10:100-8.
51. Mundinger TO, Taborsky GJ. Differential action of hepatic sympathetic neuropeptides: metabolic action of galanin, vascular action of NPY. Am J Physiol-Endoc M. 2000;278:E390-7.
52. Ren W, Hua M, Cao F, Zeng W. The Sympathetic-immune milieu in metabolic health and diseases: insights from pancreas, liver, intestine, and adipose tissues. Adv Sci. 2023;11:2306128.
53. Liu J, Bisschop PH, Eggels L, et al. Intrahypothalamic Estradiol regulates glucose metabolism via the sympathetic nervous system in female rats. Diabetes. 2013;62:435-43.
54. Liu L, Huang Z, Zhang J, et al. Hypothalamus-sympathetic-liver axis mediates the early phase of stress-induced hyperglycemia in the male mice. Nat Commun. 2024;15:8632.
55. Latour MG, Lautt W. The hepatic vagus nerve in the control of insulin sensitivity in the rat. Auton Neurosci. 2002;95:125-30.
56. Xue C, Aspelund G, Sritharan KC, Wang JP, Slezak LA, Andersen DK. Isolated hepatic cholinergic denervation impairs glucose and glycogen metabolism. J Surg Res. 2000;90:19-25.
57. Lee KC, Miller RE. The hepatic vagus nerve and the neural regulation of insulin secretion*. Endocrinology. 1985;117:307-14.
58. Moore MC, Coate KC, Winnick JJ, An Z, Cherrington AD. Regulation of hepatic glucose uptake and storage in vivo. Adv Nutr. 2012;3:286-94.
59. Perseghin G, Regalia E, Battezzati A, et al. Regulation of glucose homeostasis in humans with denervated livers. J Clin Investig. 1997;100:931-41.
60. Mahley RW, Innerarity TL, Rall SC, Weisgraber KH. Plasma lipoproteins: apolipoprotein structure and function. J Lipid Res. 1984;25:1277-94.
61. Hurr C, Simonyan H, Morgan DA, Rahmouni K, Young CN. Liver sympathetic denervation reverses obesity‐induced hepatic steatosis. J Physiol. 2019;597:4565-80.
62. Bruinstroop E, Eliveld J, Foppen E, et al. Hepatic denervation and dyslipidemia in obese Zucker (fa/fa) rats. Int J Obes. 2015;39:1655-8.
63. Carnagarin R, Tan K, Adams L, et al. Metabolic dysfunction-associated fatty liver disease (MAFLD) - a condition associated with heightened sympathetic activation. Int J Mol Sci. 2021;22:4241.
64. Raulin A, Martens YA, Bu G. Lipoproteins in the central nervous system: from biology to pathobiology. Annu Rev Biochem. 2022;91:731-59.
65. Zhu Y, Yao L, Gallo-ferraz AL, et al. Sympathetic neuropeptide Y protects from obesity by sustaining thermogenic fat. Nature. 2024;634:243-50.
66. Hackl MT, Fürnsinn C, Schuh CM, et al. Brain leptin reduces liver lipids by increasing hepatic triglyceride secretion and lowering lipogenesis. Nat Commun. 2019;10:2717.
67. Metz M, Beghini M, Wolf P, et al. Leptin increases hepatic triglyceride export via a vagal mechanism in humans. Cell Metab. 2022;34:1719-1731.e5.
68. Hwang J, Lee S, Okada J, et al. Liver-innervating vagal sensory neurons are indispensable for the development of hepatic steatosis and anxiety-like behavior in diet-induced obese mice. Nat Commun. 2025;16:991.
69. Elsing C, Hubner C, Fitscher BA, Kassner A, Stremmel W. Muscarinic acetylcholine receptor stimulation of biliary epithelial cells and its effect on bile secretion in the isolated perfused liver. Hepatology. 1997;25:804-13.
70. Friman S, Radberg G, Svanvik J. Adrenergic influence on bile secretion - an experimental study in the cat. Acta Physiol Scand. 2008;140:287-93.
71. Wang F, Lu Z, Wang X, Zhang Y. Impaired vagus function in rats suppresses bile acid synthesis in the liver by disrupting tight junctions and activating Fxr-Fgf15 signaling in the intestine. Biochem Biophys Res Commun. 2018;495:1490-6.
72. Chen C, Peng S, Chou Y, et al. Human liver afferent and efferent nerves revealed by 3-D/Airyscan super-resolution imaging. Am J Physiol-Endoc M. 2024;326:E107-23.
73. Rossen ND, Touhara KK, Castro J, et al. Population imaging of enterochromaffin cell activity reveals regulation by somatostatin. Proc. Natl. Acad. Sci. U.S.A. 2025;122:e2501525122.
74. Ohtani N, Kamiya T, Kawada N. Recent updates on the role of the gut-liver axis in the pathogenesis of NAFLD/NASH, HCC, and beyond. Hepatol Commun. 2023;7.
75. Kang JW, Vemuganti V, Kuehn JF, Ulland TK, Rey FE, Bendlin BB. Gut microbial metabolism in Alzheimer’s disease and related dementias. Neurotherapeutics. 2024;21:e00470.
76. Van Munster KN, Bergquist A, Ponsioen CY. Inflammatory bowel disease and primary sclerosing cholangitis: one disease or two? J Hepatol. 2024;80:155-68.
77. Kalaitzakis E. Gastrointestinal dysfunction in liver cirrhosis. World J Gastroenterol. 2014;20:14686.
78. Wang S, Farokhian A, Shen B. Clinical association between inflammatory bowel disease and primary sclerosing cholangitis: what changes after colectomy and liver transplantation? egastro. 2025;3:e100199.
79. Günzel D, Fromm M. Claudins and other tight junction proteins. in: prakash ys, editors. comprehensive physiology. Wiley; 2012. pp. 1819-52.
80. Bonaz B, Sinniger V, Pellissier S. Anti‐inflammatory properties of the vagus nerve: potential therapeutic implications of vagus nerve stimulation. The Journal of Physiology. 2016;594:5781-90.
81. Nijhuis LE, Olivier BJ, De Jonge WJ. Neurogenic regulation of dendritic cells in the intestine. Biochem Pharmacol. 2010;80:2002-8.
82. Nakai A, Suzuki K. Adrenergic control of lymphocyte trafficking and adaptive immune responses. Neurochem Int. 2019;130:104320.
83. Henriksen EKK, Jørgensen KK, Kaveh F, et al. Gut and liver T-cells of common clonal origin in primary sclerosing cholangitis-inflammatory bowel disease. J Hepatol. 2017;66:116-22.
84. Liang H, Xu L, Zhou C, Zhang Y, Xu M, Zhang C. Vagal activities are involved in antigen‐specific immune inflammation in the intestine. J Gastroenterol Hepatol. 2011;26:1065-71.
85. Langness S, Kojima M, Coimbra R, Eliceiri BP, Costantini TW. Enteric glia cells are critical to limiting the intestinal inflammatory response after injury. Am J Physiol - Gastrointest Liver Physiol. 2017;312:G274-82.
86. Teratani T, Mikami Y, Nakamoto N, et al. The liver-brain-gut neural arc maintains the Treg cell niche in the gut. Nature. 2020;585:591-6.
87. He X, Hu M, Xu Y, et al. The gut-brain axis underlying hepatic encephalopathy in liver cirrhosis. Nat Med. 2025;31:627-38.
88. Sun J, Debosch BJ. Reciprocal oscillation and host-microbial communication in liver metabolic control. Gut Microbes. 2025;17:2545413.
89. Zhang J, Chen K, Chen F. Exploring the impact of the liver-intestine-brain axis on brain function in non-alcoholic fatty liver disease. J Pharm Anal. 2025;15:101077.
90. Zhang T, Wang W, Li J, et al. Free fatty acid receptor 4 modulates dietary sugar preference via the gut microbiota. Nat Microbiol. 2025;10:348-61.
91. Li H, Yuan W, Tian Y, et al. Eugenol alleviated nonalcoholic fatty liver disease in rat via a gut-brain-liver axis involving glucagon-like Peptide-1. Arch Biochem Biophys. 2022;725:109269.
92. Wan M. Role of gut-brain axis dysregulation in the pathogenesis of non-alcoholic fatty liver disease: mechanisms and therapeutic implications. Am J Transl Res. 2025;17:3276-92.
93. Kwiecien S, Sliwowski Z, Pajdo R, Ptak-Belowska A, Brzozowski T. Role of brain-gut axis in mechanism of gastrointestinal defense. J Physiol Pharmacol. 2025;76:487-99.
94. Raybould HE. Nutrient sensing in the gastrointestinal tract: possible role for nutrient transporters. J Physiol Biochem. 2008;64:349-56.
95. Secher A, Jelsing J, Baquero AF, et al. The arcuate nucleus mediates GLP-1 receptor agonist liraglutide-dependent weight loss. J Clin Investig. 2014;124:4473-88.
96. Dai F, Yin J, Chen JDZ. Effects and mechanisms of vagal nerve stimulation on body weight in diet-induced obese rats. Obes Surg. 2020;30:948-56.
97. Krieger J, Arnold M, Pettersen KG, Lossel P, Langhans W, Lee SJ. Knockdown of GLP-1 receptors in vagal afferents affects normal food intake and glycemia. Diabetes. 2016;65:34-43.
98. Hoffman S, Alvares D, Adeli K. GLP-1 attenuates intestinal fat absorption and chylomicron production via vagal afferent nerves originating in the portal vein. Molecular Metabolism. 2022;65:101590.
99. Visschers RG, Luyer MD, Schaap FG, Olde Damink SW, Soeters PB. The gut-liver axis. Curr Opin Clin Nutr Metab Care. 2013;16:576-81.
100. Muller PA, Matheis F, Schneeberger M, Kerner Z, Jové V, Mucida D. Microbiota-modulated CART+ enteric neurons autonomously regulate blood glucose. Science. 2020;370:314-21.
101. Macpherson AJ, Pachnis V, Prinz M. Boundaries and integration between microbiota, the nervous system, and immunity. Immunity. 2023;56:1712-26.
102. Pan L, Xie L, Yang W, et al. The role of brain-liver-gut axis in neurological disorders. Burns & Trauma. 2025;13:tkaf011.
103. Chen W, Deng F, Tsai Y. Lactiplantibacillus plantarum as a psychobiotic strategy targeting parkinson’s disease: a review and mechanistic insights. Nutrients. 2025;17:3047.
104. Ren W, Chen J, Wang W, et al. Sympathetic nerve-enteroendocrine L cell communication modulates GLP-1 release, brain glucose utilization, and cognitive function. Neuron. 2024;112:972-990.e8.
105. Cook TM, Gavini CK, Jesse J, et al. Vagal neuron expression of the microbiota-derived metabolite receptor, free fatty acid receptor (FFAR3), is necessary for normal feeding behavior. Mol Metab. 2021;54:101350.
106. Sharma D, Malik A, Locher V, et al. A myeloid Tet2-IL-1β axis modulates intestinal inflammation by restricting catecholaminergic stimulation of enterochromaffin cell differentiation. Immunity. 2025;58:2785-2798.e4.
107. Shen B, Gu T, Shen Z, et al. Escherichia coli promotes endothelial to mesenchymal transformation of liver sinusoidal endothelial cells and exacerbates nonalcoholic fatty liver disease via its flagellin. Cell Mol Gastroenterol Hepatol. 2023;16:857-79.
108. Ahmed O, Caravaca AS, Crespo M, et al. Hepatic stellate cell activation markers are regulated by the vagus nerve in systemic inflammation. Bioelectron Med. 2023;9:6.
109. Boicean A, Birlutiu V, Ichim C, Brusnic O, Onișor DM. Fecal microbiota transplantation in liver cirrhosis. Biomedicines. 2023;11:2930.
110. Wu L, Li J, Zou J, Tang D, Chen R. Vagus nerve modulates acute-on-chronic liver failure progression via CXCL9. Chin Med J (Engl). 2024;138:1103-15.
111. Siripen N, Chonprasertsuk S, Aumpan N, Siramolpiwat S. Efficacy of a 12-week supervised home-based exercise program and nutritional supplementation in cirrhotic patients with sarcopenia: A prospective pilot study. Clinical Nutrition ESPEN. 2026;72:102810.
112. Carabotti M, Scirocco A, Maselli M A, Severi C. The gut-brain axis: interactions between enteric microbiota, central and enteric nervous systems. Ann Gastroenterol. 2015;28:203-9.
113. Gershon MD, Margolis KG. The gut, its microbiome, and the brain: connections and communications. J Clin Investig. 2021;131:e143768.
114. Bonaz B, Sinniger V, Pellissier S. Vagus nerve stimulation: a new promising therapeutic tool in inflammatory bowel disease. J Intern Med. 2017;282:46-63.
115. Xie Z, Zhang X, Zhao M, et al. The gut-to-brain axis for toxin-induced defensive responses. Cell. 2022;185:4298-4316.e21.
116. Coulter AA, Rebello CJ, Greenway FL. Centrally acting agents for obesity: past, present, and future. Drugs. 2018;78:1113-32.
117. Ma L, Wang H, Hashimoto K. The vagus nerve: an old but new player in brain-body communication. Brain, Behavior, and Immunity. 2025;124:28-39.
118. Patterson TT, Nicholson S, Wallace D, Hawryluk GW, Grandhi R. Complex feed-forward and feedback mechanisms underlie the relationship between traumatic brain injury and the gut-microbiota-brain axis. Shock. 2019;52:318-25.
119. Yang Y, Eguchi A, Wan X, Mori C, Hashimoto K. Depression-like phenotypes in mice with hepatic ischemia/reperfusion injury: A role of gut-microbiota-liver-brain axis via vagus nerve. J Affect Disord. 2024;345:157-67.
120. Yang X, Qiu K, Jiang Y, Huang Y, Zhang Y, Liao Y. Metabolic crosstalk between liver and brain: from diseases to mechanisms. Int J Mol Sci. 2024;25:7621.
121. Yin J. Transcutaneous vagal nerve stimulation for gastrointestinal disorders. J Transl Gastroenterol. 2025;000:000.
122. Hong GS, Zillekens A, Schneiker B, et al. Non‐invasive transcutaneous auricular vagus nerve stimulation prevents postoperative ileus and endotoxemia in mice. Neurogastroenterology Motil. 2018;31:e13501.
123. Jin H, Guo J, Liu J, et al. Anti-inflammatory effects and mechanisms of vagal nerve stimulation combined with electroacupuncture in a rodent model of TNBS-induced colitis. Am J Physiol - Gastrointest Liver Physiol. 2017;313:G192-202.
124. Sahn B, Pascuma K, Kohn N, Tracey KJ, Markowitz JF. Transcutaneous auricular vagus nerve stimulation attenuates inflammatory bowel disease in children: a proof-of-concept clinical trial. Bioelectron Med. 2023;9:23.
125. Brinkman DJ, Simon T, Ten Hove AS, et al. Electrical stimulation of the splenic nerve bundle ameliorates dextran sulfate sodium-induced colitis in mice. J Neuroinflammation. 2022;19:155.
126. Zhu Y, Xu F, Lu D, et al. Transcutaneous auricular vagal nerve stimulation improves functional dyspepsia by enhancing vagal efferent activity. Am J Physiol - Gastrointest Liver Physiol. 2021;320:G700-11.
127. Shi X, Hu Y, Zhang B, Li W, Chen JD, Liu F. Ameliorating effects and mechanisms of transcutaneous auricular vagal nerve stimulation on abdominal pain and constipation. JCI Insight. 2021;6:e150052.
128. Yuan P, Taché Y. Abdominal surgery induced gastric ileus and activation of M1-like macrophages in the gastric myenteric plexus: prevention by central vagal activation in rats. Am J Physiol - Gastrointest Liver Physiol. 2017;313:G320-9.
129. Jiang Y, Wu L, Zhu X, Bian H, Gao X, Xia M. Advances in management of metabolic dysfunction-associated steatotic liver disease: from mechanisms to therapeutics. Lipids Health Dis. 2024;23:95.
130. Liu A, Huang M, Xi Y, Deng X, Xu K. Orchestration of gut-liver-associated transcription factors in MAFLD: from cross-organ interactions to therapeutic innovation. Biomedicines. 2025;13:1422.
131. John S, Bhowmick K, Park A, Huang H, Yang X, Mishra L. Recent advances in targeting obesity, with a focus on TGF-β signaling and vagus nerve innervation. Bioelectron Med. 2025;11:10.
132. Ueda T, Tategaki A, Hamada K, et al. Effects of Pediococcus acidilactici R037 on serum triglyceride levels in mice and rats after oral administration. J Nutr Sci Vitaminol. 2018;64:41-7.
133. Feng R, Yang W, Feng W, et al. Time-restricted feeding ameliorates non-alcoholic fatty liver disease through modulating hepatic nicotinamide metabolism via gut microbiota remodeling. Gut Microbes. 2024;16:2390164.
134. Gupta H, Suk KT, Kim DJ. Gut microbiota at the intersection of alcohol, brain, and the liver. JCM. 2021;10:541.
135. Pan Y, Yang Y, Wu J, Zhou H, Yang C. Efficacy of probiotics, prebiotics, and synbiotics on liver enzymes, lipid profiles, and inflammation in patients with non-alcoholic fatty liver disease: a systematic review and meta-analysis of randomized controlled trials. BMC Gastroenterol. 2024;24:283.
136. Kobayashi H, Iida T, Ochiai Y, et al. Neuro-mesenchymal interaction mediated by a β2-adrenergic nerve growth factor feedforward loop promotes colorectal cancer progression. Cancer Discovery. 2025;15:202-26.
137. Wang T, Teng B, Yao DR, Gao W, Oka Y. Organ-specific sympathetic innervation defines visceral functions. Nature. 2024;637:895-902.
138. Shi X, Zhao L, Luo H, et al. Transcutaneous auricular vagal nerve stimulation is effective for the treatment of functional dyspepsia: a multicenter, randomized controlled study. Am J Gastroenterol. 2024;119:521-31.
139. Kovacic K, Hainsworth K, Sood M, et al. Neurostimulation for abdominal pain-related functional gastrointestinal disorders in adolescents: a randomised, double-blind, sham-controlled trial. Lancet Gastroenterol Hepatol. 2017;2:727-37.
140. Krasaelap A, Sood MR, Li B, et al. Efficacy of auricular neurostimulation in adolescents with irritable bowel syndrome in a randomized, double-blind Trial. Clin Gastroenterol Hepatol. 2020;18:1987-1994.e2.
141. Huang Z, Lin Z, Lin C, et al. Transcutaneous electrical acustimulation improves irritable bowel syndrome with constipation by accelerating colon transit and reducing rectal sensation using autonomic mechanisms. Am J Gastroenterol. 2022;117:1491-501.
142. Zhou J, Wang J, Ning B, et al. Sustained ameliorating effects and autonomic mechanisms of transcutaneous electrical acustimulation at ST36 in patients with chronic constipation. Front Neurosci. 2022;16:1038922.
143. Liu Z, Yan S, Wu J, et al. Acupuncture for chronic severe functional constipation: a randomized trial. Ann Intern Med. 2016;165:761-9.
144. Liu B, Wu J, Yan S, et al. Electroacupuncture vs prucalopride for severe chronic constipation: a multicenter, randomized, controlled, noninferiority trial. Am J Gastroenterol. 2021;116:1024-35.
145. Iqbal F, Thomas GP, Tan E, et al. Transcutaneous sacral electrical stimulation for chronic functional constipation. Diseases of the Colon & Rectum. 2016;59:132-9.
146. Iqbal F, Collins B, Thomas GP, et al. Bilateral transcutaneous tibial nerve stimulation for chronic constipation. Colorectal Dis. 2016;18:173-8.
147. Gokce AH, Gokce FS. Effects of bilateral transcutaneous tibial nerve stimulation on constipation severity in geriatric patients: a prospective clinical study. Geriatrics Gerontology Int. 2019;20:101-5.
148. Chase J, Robertson VJ, Southwell B, Hutson J, Gibb S. Pilot study using transcutaneous electrical stimulation (interferential current) to treat chronic treatment‐resistant constipation and soiling in children. J Gastroenterol Hepatol. 2005;20:1054-61.
149. Clarke MC, Chase JW, Gibb S, et al. Decreased colonic transit time after transcutaneous interferential electrical stimulation in children with slow transit constipation. J Pediatr Surg. 2009;44:408-12.
150. Clarke MC, Chase JW, Gibb S, Hutson JM, Southwell BR. Improvement of quality of life in children with slow transit constipation after treatment with transcutaneous electrical stimulation. J Pediatr Surg. 2009;44:1268-73.
151. Ismail KA, Chase J, Gibb S, et al. Daily transabdominal electrical stimulation at home increased defecation in children with slow-transit constipation: a pilot study. J Pediatr Surg. 2009;44:2388-92.
152. Yik YI, Ismail KA, Hutson JM, Southwell BR. Home transcutaneous electrical stimulation to treat children with slow-transit constipation. J Pediatr Surg. 2012;47:1285-90.
153. Queralto M, Vitton V, Bouvier M, Abysique A, Portier G. Interferential therapy: a new treatment for slow transit constipation. A pilot study in adults. Colorectal Dis. 2012;15.
154. Clarke MC, Catto-smith AG, King SK, et al. Transabdominal electrical stimulation increases colonic propagating pressure waves in paediatric slow transit constipation. J Pediatr Surg. 2012;47:2279-84.
155. Yang Y, Yim J, Choi W, Lee S. Improving slow-transit constipation with transcutaneous electrical stimulation in women: A randomized, comparative study. Women & Health. 2016;57:494-507.
156. Moore JS, Gibson PR, Burgell RE. Randomised clinical trial: transabdominal interferential electrical stimulation vs sham stimulation in women with functional constipation. Aliment Pharmacol Ther. 2020;51:760-9.
157. Xiao W, Liu Y. Rectal hypersensitivity reduced by acupoint TENS in patients with diarrhea-predominant irritable bowel syndrome: a pilot study. Dig Dis Sci. 2004;49:312-9.
158. Xiao Y, Xu F, Lin L, Chen JD. Transcutaneous Electrical Acustimulation Improves Constipation by Enhancing Rectal Sensation in Patients With Functional Constipation and Lack of Rectal Sensation. Clin Transl Gastroenterol. 2022;13:e00485.
159. Çoban Ş, Akbal E, Köklü S, et al. Clinical trial: transcutaneous interferential electrical stimulation in individuals with irritable bowel syndrome - a prospective double-blind randomized study. Digestion. 2012;86:86-93.
160. Adams D. The course and prognostic factors of familial amyloid polyneuropathy after liver transplantation. Brain. 2000;123:1495-504.
161. Zhao J, Zhao X, Wang Q, et al. Efficacy and safety of electroacupuncture for metabolic dysfunction-associated fatty liver disease: a study protocol for a multicentre, randomised, sham acupuncture-controlled, patient-blinded clinical trial. BMJ Open. 2024;14:e084768.
162. Fu L, Huang L, Gao Y, et al. Investigating the efficacy of acupuncture in treating patients with metabolic-associated fatty liver disease: a protocol for a randomised controlled clinical trial. BMJ Open. 2024;14:e081293.
Cite This Article
How to Cite
Download Citation
Export Citation File:
Type of Import
Tips on Downloading Citation
Citation Manager File Format
Type of Import
Direct Import: When the Direct Import option is selected (the default state), a dialogue box will give you the option to Save or Open the downloaded citation data. Choosing Open will either launch your citation manager or give you a choice of applications with which to use the metadata. The Save option saves the file locally for later use.
Indirect Import: When the Indirect Import option is selected, the metadata is displayed and may be copied and pasted as needed.
About This Article
Copyright

Data & Comments
Data
0













Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at [email protected].